SCOTCAT.COM
your internet guide to all things catfish
| Mystus impluviatus. A New Species of Bagrid Catfish (Teleostei: Bagridae) from Eastern Borneo | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
by Heok Hee Ng |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
THE
genus Mystus Scopoli, 1777 is a group of
bagrid catfishes found in streams and rivers throughout
South and Southeast Asia. The genus Macrones
was used by earlier authors, but Macrones
Duméril, 1856, is preoccupied by Macrones
Newman, 1841, in Coleoptera and Mystus Scopoli,
1777, was revived by Fowler (1928) as a replacement
name. Mystus is poorly diagnosed and is polyphyletic
(Mo, 1991). Twentytwo of the 45 nominal species of
Mystus are found in Southeast Asia, 19 of
which are currently considered valid. DESCRIPTION Holotype.—CAS 97049, male, 87.0 mm SL; Borneo: Kalimantan Timur, small river on road 12.2 km from Sebulu crossroads towards forest and 59 km from Air Putih crossroads (08169S, 117809E); M. S. Christensen, H. Christensen, and H. Aschanuddin, 31 July 1982. Paratypes.—CAS 94621, 2 juveniles, 20.3–22.8 mm SL; Borneo: Kalimantan Timur, beneath floating water hyacinth island about 6 m2, flooded swamp forest ca. 6 km SE of Kota Bangun (0°16'S, 116°34'E); M. S. Christensen et al., 1983. CAS 95038, 4 males, 76.2–119.6 mm SL, 1 female 80.8 mm SL; USNM 371267, 1 male, 83.0 mm SL, 1 female, 81.1 mm SL; Borneo: Kalimantan Timur, remnant pool in swamp forest, approximately 120 m2 along Sungai Kedang Kepala, ca. 15 km NNW of Muara Kaman (0°9'S, 116°43,E); M. S. Christensen and M. Schellenberg, November 1982. CAS 95035, 1 male, 98.9 mm SL; 1 female, 113.0 mm SL; MZB 10703, 1 female, 96.1 mm SL; Borneo: Kalimantan Timur, Belayan River (0°7'N, 116°22'E); M. S. Christensen et al., 10 August 1982. CAS 97045, 3 juveniles, 25.7–30.0 mm SL; Borneo: Kalimantan Timur, from small stream on logging trail going inland from southern junction of Sebulu, side road to stream approximately 4 km (0°16'S,117°0'E); M.S. Christensen and G.von Drabich, 31 October 1982.
Fig. 1. Mystus impluviatus, paratype male, CAS 95035, 98.9 mm SL.
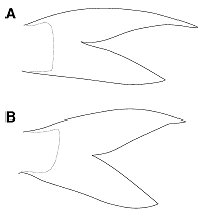
Diagnosis.—Mystus impluviatus differs from both M. castaneus and M. nigriceps in having a second posterior fontanel on the supraoccipital (Fig. 2). It further differs from M. castaneus in having a shallower adipose fin (4.7–6.0% SL vs 6.1– 7.0), smaller serrations on the posterior edge of the pectoral spine (maximum height of serrations 3.6–4.8% of spine length vs 6.9–8.0), and a more deeply forked caudal fin with a more slender upper lobe (Fig. 3). Mystus impluviatus further differs from M. nigriceps in having a longer adipose fin base (31.6–36.1% SL vs 26.3– 31.4), longer second and third dorsal fin rays, causing the dorsal fin to appear less rounded, and a brown body with a distinct dark triangular patch on the base of the caudal peduncle (vs greenish gray body with a diffuse dark triangular patch on the base of the caudal peduncle).
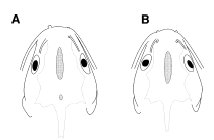
Fig. 2. Schematic illustration of dorsal views of heads of (A) Mystus impluviatus, CAS 97049, holotype, 87.0 mm SL; (B) Mystus castaneus and Mystus nigriceps (M. castaneus, UMMZ 155714, 77.9 mm SL illustrated), showing presence of second posterior fontanel on supraoccipital in M. impluviatus. Scale bar represents 10 mm. Description.—Head depressed; dorsal profile slightly convex and ventral profile almost straight. Bony elements of dorsal surface of head covered with thin skin; bones readily visible, ornamented with numerous fine grooves. Eye ovoid, horizontal axis longest, with free margin; located entirely in dorsal half of head. Gill openings wide, extending from posttemporal to beyond isthmus. Gill membranes free from isthmus, with 9 (n 5 11) or 10 (n 5 4) branchiostegal rays. First branchial arch with 6 + 14 (n = 4), 5 + 20 (n = 2), 5 + 21 (n = 2), 6 + 19 (n = 1), 6 + 20 (n = 4) or 6 + 21 (n = 2) gill rakers. Mouth subterminal, fleshy upper lip extending anteriorly beyond upper jaw. Oral teeth small and viliform, in irregular rows on all tooth-bearing surfaces. Premaxillary tooth band rounded, of equal width throughout. Dentary tooth band much narrower than premaxillary tooth band at symphysis, tapering laterally. Vomerine tooth band unpaired, continuous across midline; smoothly arched along anterior margin, tapering laterally to point extending posteriorly well past level of premaxillary band; band width narrower than premaxillary band at midline, widening laterally and then tapering to a sharp point posterolaterally.
Fig. 3. Schematic illustration of caudal fins of (A) Mystus impluviatus, CAS 95038, paratype, 83.0 mm SL; (B) Mystus castaneus, UMMZ 155714, 77.9 mm SL. Scale bar represents 5 mm.
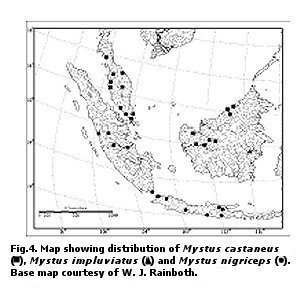
Fig. 4. Map showing distribution of Mystus castaneus, Mystus impluviatus and Mystus nigriceps (c). Base map courtesy of W. J. Rainboth. Barbels in four pairs. Maxillary barbel long and slender, extending beyond caudal fin. Nasal barbel slender, extending to halfway between posterior orbital margin and dorsalmost extent of gill opening. Inner mandibular-barbel origin close to midline; barbel thicker and longer than nasal barbel and extending just beyond base of last pectoral fin ray. Outer mandibular barbel originates posterolateral of inner mandibular barbel, extending to base of first pelvic fin ray. Body slightly
compressed, becoming more so toward caudal peduncle.
Dorsal profile rising evenly but not steeply from
tip of snout to origin of dorsal fin and sloping gently
ventrally from origin of dorsal fin to end of caudal
peduncle. Ventral profile slightly convex to anal
fin base, then sloping slightly dorsally to end of
caudal peduncle. Dorsal fin with spinelet, spine, and 6 (n = 2) or 7 (n = 13) rays. Origin of dorsal fin slightly anterior to midline. Dorsal fin margin convex, usually with anterior branch of fin rays longer than other branches. Dorsal fin spine short, straight and slender, posterior edge with low irregular serrations. Pectoral fin
with stout spine, sharply pointed at tip, and 8 (n
= 4), 9 (n = 2) or 9,i (n = 9) Pelvic fin origin at vertical through posterior end of dorsal-fin base, with i,5 (n = 15) rays and slightly convex margin; tip of adpressed fin not reaching anal fin origin. Anus and urogenital openings located at vertical through middle of adpressed pelvic fin. Males with a long, slender genital papilla extending to base of second anal fin ray. Adipose fin with convex margin for entire length; anterior end reaching to base of last dorsal fin ray. Anal fin base ventral to posterior half of adipose fin, with iv,7 (n = 2), iv,8 (n = 9) or iv,9 (n = 4) rays and curved posterior margin. Caudal peduncle moderately slender. Caudal fin deeply forked, with i,7,7,i (n = 2) or i,7,8,i (n = 13) principal rays; upper lobe slender and lanceolate, lower lobe pointed. Procurrent rays extend only slightly anterior to fin base. Morphometric data as in Table 1.
TABLE 1. MORPHOMETRIC DATA FOR Mystus impluviatus (n = 10). Range (mean ± SD) % SL
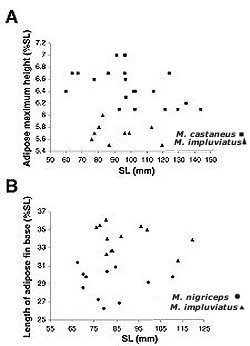
Fig. 5. (A) Maximum height of the adipose fin (%SL) plotted against SL (mm) for Mystus castaneus and Mystus impluviatus; (B) Length of the adipose fin base (%SL) plotted against SL (mm) for M. impluviatus and Mystus nigriceps. Color in alcohol.—Dorsal surface of head and body uniform medium brown, with a very faint dark gray humeral spot and a distinct oval or triangular dark gray spot on base of caudal peduncle.Ventral surfaces of head and body dirty white; adipose fin and fin rays of all fins brown; interradial membranes of all fins with scattered melanophores. Dorsal half of barbels brown at base, gradually turning to dark yellow on ventral half and tips. Distribution.—Known from the Mahakam River drainage in eastern Borneo (Fig. 4). Etymology.—From
the Latin impluvium, which refers to a skylight; in
allusion to the second posterior DISCUSSION The difference in the height of the adipose fin between M. castaneus and M. impluviatus cannot be explained by ontogenetic change alone, as a plot of the maximum height of the adipose fin against SL (Fig. 5a) shows. A plot of the length of the adipose fin base against SL (Fig. 5b) also shows that ontogeny alone cannot explain the differences observed between M. nigriceps and M. impluviatus. Lines drawn through the plot of all data from both species as a single series have very low r2-values (0.0002 for the maximum height of the adipose fin and 0.0296 for the length of the adipose fin base). Mystus impluviatus appears to be endemic to the Mahakam River drainage (or, at least, to eastern Borneo). This is not surprising, given that the Mahakam River was isolated from the other main Southeast Asian river drainages throughout the low sea level periods of the Pleistocene (Voris, 2000). Although the endemism of the Mahakam fish fauna is currently thought to be low (about 9%; Kottelat, 1994), recent studies have identified at least three more endemic species of catfishes (e.g., Ng, 2000, 2001; Ng and Ng, 2001). Therefore, the fish fauna of the Mahakam River deserves further study as it is likely that there are more endemic species in the river system. COMPARATIVE
MATERIAL Mystus castaneus:
ZRC 41848, holotype, 121.4 mm SL; Borneo: Sarawak,
Serian market, from Sungai Sadong. ZRC 28176, 1 paratype,
77.6 mm SL; Malaysia: Johor, Kota Tinggi, Sungai Mupor.
ZRC 39419, 3 paratypes, 124.0–134.5 mm SL; Sarawk:
Serian market. ZRC 40487, 8 paratypes, 86.5–114.1
mm SL; Borneo: Brunei, Tutong district, Sungai Merimbun,
outflow from Tasik Merimbun draining into Sungai Tutong.
ZRC 40490, 6 paratypes, 92.7–143.8 mm SL; Borneo:
Sarawak, Serian market. UMMZ 155689, 1 ex., 101.9
mm SL; UMMZ 155713, 1 ex., 58.7 mm SL; UMMZ 155714,
6 ex., 38.8– 87.8 mm SL; Sumatra: Musi River,
Muara Klingi. LITERATURE
CITED |


If you would like to contribute an article, please e-mail me. You will of course be credited for your work.
If you would like to donate any denomination of money to the site just click the above link button. All proceeds will go to running the site and hopefully to keep it going for a few years yet.
Print or e-mail this factsheet below
