SCOTCAT.COM
your internet guide to all things catfish
| The striped catfishes of the genus Mystus Scopoli, 1777 (Siluriformes: Bagridae) |
| by Steven Grant |
This
article has been written for interested aquarists (or,
to a much lesser degree, ichthyologists), to contribute
towards resolving the identity and validity of the striped
bagrid catfishes of the genus Mystus Scopoli,
1777.
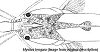
Roberts restricted Mystus to the species with long and high adipose fins, long maxillary barbels, deeply forked caudal fins (with upper lobe being longer) and with the split fontanel. He did not propose any generic name(s) for the remaining species. Jayaram & Sanyal (2003) felt that more of the species could be included in the sensu stricto group to the ones included by Roberts, and felt that some of his characteristics were unreliable. There is a generic name that already exists which could be resurrected in the future for some of the remaining species: Aspidobagrus Bleeker, 1862. Recent authors have mentioned Hypselobagrus Bleeker, 1862, but if one follows Roberts (1992) work on the synonymy of its type species, this genus would be a synonym of Mystus. Looking at the morphology of what is currently considered to be the type species of Mystus, I personally doubt how it can be included as belonging to the same morphological group as the species Roberts (1992) lists in sensu stricto. Basically it is still a bit of a mess and I think that for the time being they will have to be referred to as Mystus, but I wouldn’t bet against Hypselobagrus and/or Aspidobagrus being resurrected. With all the species I have tried to get images of the preserved type specimens and/or reproduced the original drawing if one was provided. The reason I have done this is that I have found that unless one refers to the type specimens or drawings of them, one cannot be 100% sure that you are talking about the species in question. I appreciate that for some people images or drawings of dead, sometimes now colourless, fish may not be the best way to assist in identification but misidentifications have been made previously and I feel this is the safest way. However, even with type material to refer to, some of the multi-striped species are still difficult to tell apart when looking at live fish, and sometimes one has to use the morphology of the fontanel (which is not easy in live fish) and supra-occipital process (a bony extension of the skull which extends towards the ‘basal bones’ of the dorsal fin – see image below).
Sometimes the length, placement and height of the adipose fin in combination with maxillary barbel length and body pattern are a good defining character. Roberts (1992) separates the ‘shoulder spot’ into two types. In some species it is a spot or a dark area, whereas in some species it is a semi-ocellus as it has a dark or black spot with a pale or bright imperfect ring around it: Humeral
semi-ocellus / spot Tympanic semi-ocellus
/ spot But for some species the only definite differences are that of measurements and proportions (morphometrics) and also counts of gill rakers, vertebrae etc (meristics). Obviously I have not mentioned these differences as they are useless for most users of this article. I hope this article will go some short way in assisting anyone interested in the identification of this ‘group’ of catfish. Mystus albolineatus
Roberts, 1994
(Please see
image of live specimen)
See holotype CAS 79030 which originated from Prachinburi Market, Bangpakong basin, Thailand, but the species is also present in the lower Mekong basin, Cambodia. Largest type specimen is 13.5 cm SL. See also a paratype (ANSP 16453), and a live specimen by Kamphol.This species has a white or pale stripe running along the lateral line. The stripe is bordered above and below by dark patterning. Some specimens have a dark spot at the dorsal fin base; some a small dark triangular spot at the middle of the body near the base of the caudal fin (midpeduncular spot); some with a humeral semi-ocellus.Can be visually distinguished from other species by combination of a very long adipose fin which originates more or less directly after the dorsal fin, very long maxillary barbels which extend at least to the posterior point of the body, the white lateral streak, and the caudal fin lobes curling inwards. Mystus ankutta Pethiyagoda, Silva, & Maduwage, 2008
See image of one of the type specimens. This species is only present in south-western Sri Lanka, from the Kelani, Kalu, Bentara, and Nilwada Rivers. The largest type specimen is 7.92 cm SL.This species has a thin black stripe running along the lateral line of the body, a diffuse grey humeral blotch and a distinct black sub-triangular blotch on the hypural (near the caudal peduncle) base.Can be distinguished from M. canarensis by the supra-occipital process being longer (extending further towards the nuchal plate). It also appears to be less elongated than M. canarensis. It is very similar to the true M. malabaricus; I haven’t used the detailed information in Pethiyagoda et al, 2008 because I respectfully do not agree on the identity of M. malabaricus or M. armatus used therein (see further discussion under M. canarensis, M. malabaricus, and M. oculatus). In my opinion the specimens used in Pethiyagoda et al as M. armatus (apart from the photograph of the syntype of M. armatus) are probably closer to M. malabaricus, although they slightly differ to AMS B.7573 and NMW 45165 which I use here as representing M. malabaricus. Using the aforementioned specimens, M. ankutta can be distinguished from M. malabaricus by the narrower anterior portion of the anterior fontanel.It differs from M. keletius by the fontanel being split (vs not split) by an epiphyseal bar. It differs from M. oculatus (and its proable junior synonym M. armatus) by the posterior portion of the fontanel not extending near the base of the supra-occipital process (versus extending). Mystus atrifasciatus Fowler, 1937 (Please see images of live specimens)
(Click thumbnail for image)
(Click thumbnail for image)
See holotype ANSP 67907 (which originates from Pitsanulok, Thailand) and paratype ANSP 67908, and live specimen by Kamphol. Largest type specimen is 8.62 cm SL but is believed to grow to at least 11 cm SL. Colouration as per original description: “Back and upper surface of head brown. Dark to blackish grey median lateral band, wide as vertical eye diameter and including lateral line, bounded above by whitish parallel longitudinal narrower band its whole extent, and below by whitish colour of under surfaces of body. Pale brownish streak, narrowing behind, back from pectoral axil until over front of anal. Iris grey. Lips pale or whitish. All barbels pale, with brown margins and nasal and maxillary pairs darker. Fins all more or less dull brownish.” Mystus bleekeri
(Day, 1877)
See paralectotype AMS B.7999 which supposedly originates from Seharunpore/ Seeharanpore/ Sethrampoor, West Bengal State which now appears to be a suburb of Calcutta, India. And also specimen CAS 93966.Colour from original description: “Brownish grey, with two longitudinal bands, one above the other below the lateral line, some specimens have a dark shoulder spot and a dark band along the middle of the anal fin. The fins are most darkest at their edges.” The “dark shoulder spot” is a tympanic spot. Said to attain 12.5 to 13.5 cm SL. Similar to Mystus atrifasciatus but M. bleekeri has shorter maxillary barbels (in M. atrifasciatus they extend to or past the caudal peduncle), the posterior edge of the adipose fin is angled off (versus rounded in M. atrifasciatus), and has the fontanel indented (versus not). Also similar to M. vittatus (Bloch, 1794) and often mistaken for it; see under that species for information. Could at first glance be confused with M. montanus but that species has a smaller adipose fin and the fontanel is shorter, only extending to midway between eye and base of supra-occipital process. Mystus bocourti
(Bleeker, 1864)
(Please
see image of live specimen)
M. bocourti was originally described from the Mé-Nam River at Bangkok, Thailand; it is also present in Laos and Cambodia. It reaches up to 24 cm (SL). See holotype (MNHN 1553). It is sometimes still listed in the genus Heterobagrus Bleeker, 1864, although this is currently considered to be a synonym of Mystus. This species is usually uniform silver, bronze or platinum in colour but I have seen at least one specimen that has two light bands above and below the dark lateral line (Burgess, 1989, Plate 4). As per Roberts (1994) it also sometimes has a humeral semi-ocellus and / or a dark spot on the body near the base of the dorsal fin spine. It is easily differentiated from the other species by the extraordinary long dorsal fin. Mystus canarensis Grant, 1999 (Please see images of live specimens) |
|
|

(Click thumbnail for image)  (Click
thumbnail for image)
(Click
thumbnail for image) |
|
|
 (Click thumbnail for image)
(Click thumbnail for image)
|
The type specimen (see discussion below) of M. canarensis (AMS B.7624) measures 11.1 cm SL and originates from the district of Canara (or Kanara), Karnataka State, India and is probably from Mangalore (see Ferraris et al, 2000 for information on another AMS Day specimen listed as Macrones armatus - but not representing that species in my opinion - also listed as being from ‘Canara’). The coordinates given in my description were taken from the specimen label but must be in error as the coordinates given are much further north and away from the Canara region. Although Mangalore is in Day’s view of what is meant by Malabar, in the description of Hara malabarica he mentions receiving some specimens from Travancore. Travancore is still in the Malabar region but is further south than Mangalore. However, it is not clear if all the syntypes were from Travancore, and in any case Jayaram & Sanyal (2003:71) found M. canarensis specimens in Travancore (under the account of M. malabaricus), and AMS B.7624 was labelled as Hara malabarica by Day himself. It is probable that AMS B.7624 is the specimen figured by Day (1877, Plate CI fig. 2) as “Macrones Malabaricus” (but it is important to note that Day captioned it as his species, not Jerdon’s Bagrus malabaricus), showing a fish with a tympanic semi-ocellus, a dark stripe along the lateral line, and spots on the fins making the upper portion of the dorsal fin and lower third of the anal fin appear blackish. The original drawing of Hara malabarica is not a good one. In fact in the copy that I was originally provided with the adipose fin wasn’t even showing. However, it does show in the reprint that I now have. In my opinion Fig. 12 in Jayaram & Sanyal (2003), and Fig. 119 in Easa & Shaji (2003) show M. canarensis, captioned as M. malabaricus. Please ignore the information in the description about the number of gill rakers as this is a mistake. Ferraris (2007) does not accept my neotype designation. He considers that the absence of types and nomenclatural need was not clearly demonstrated. Whitehead & Talwar (1976) found no types (which included BMNH, NMW and ZSI – where all the purported syntypes are), and I could not trace them in the BMNH. Jayaram & Sanyal class NMW 45228 as a “topotype” of Hara malabarica. Thanks to Helmut Wellendorf I have had access to images of the NMW specimen (see link above). It seems too large to be a syntype (it measures approx. 12cm SL and 14.5cm TL and Day states that his specimens from Travancore were “rather less than 4 inches in length”). Also I have been informed by Helmut that the locality label on the specimen is “Canara”. This again appears to match that of AMS.B 7624 and is probably from the same batch; I do not consider the NMW specimen to be a syntype of Hara malabarica. It is topotypical with M. canarensis and matches that species. With hindsight I should have been more specific with the information rather than just use the bibliographic reference via Eschmeyer et al to searches by other authors. The identity of Day’s species is not straight forward. It was severely wound up in a problematic identification problem with Mystus malabaricus (Jerdon, 1849) and as such the identity of the species becomes very difficult and confusing if not based on an actual type specimen. Therefore I consider the neotype to be nomenclaturally and taxonomically required, and despite the partially valid criticism of the way in which the search for the types was expressed in the designation I consider it to be valid as it meets all the requirements of Article 75 of the ICZN. It gives a base to work from to compare it to Jerdon’s M. malabaricus and M. montanus and M. armatus (sensu Day, 1865) and M. oculatus from the same region. It is my opinion that M.
armatus is possibly a synonym of M. oculatus
(see under that species for a further discussion). However,
it is clear that there are at least 4 species in the
‘Malabar’ region: Therefore when comparing M. malabaricus (using specimens under the account of M. armatus in Jayaram & Sanyal, and Pethiyagoda et al, and also AMS B.7573 and NMW 45165) it is clear that M. malabaricus and M. canarensis differ. M. canarensis has a much shorter supra-occipital process, a longer more slender body, the fontanel not reaching base of supra-occipital process (vs. almost reaching), different shaped teeth bands, and other morphometric differences. This mainly revolves around my view that Day (in his 1877 account) used a specimen not conspecific with his original type specimens, and this has led Jayaram and all others since to base what they consider M. armatus to be on a misidentification (see further discussion under M. oculatus). What would finally resolve the issue of whether M. canarensis is a different species to the true M. malabaricus is a neotype designation for Jerdon’s species. However, the identity of M. armatus must be resolved first as discussed under M. oculatus. M. canarensis is also similar at first glance to Mystus dibrugarensis and Mystus rufescens (Vinciguerra, 1890) - see under species headings for differences. See images of a very small specimen of what appears to be M. canarensis (photographed by Rahul Kumar). This specimen was caught in leaf litter in a hill stream in Wayanad, a couple of hundred miles south of Canara district. In the first picture it measures around 2 inches Total Length, in the other images it is around 4 inches TL. Compare with the images under M. malabaricus. Mystus carcio (Hamilton, 1822) (Please see image of live specimen)
This species is a dwarf one, only reaching approx. 5cm SL in captivity. It can be differentiated from all others by its small adult size, very small adipose fin, a dark horizontal mark across the tympanum, greenish yellow colouration, large laterally placed eyes, and relatively long and wide fontanel which is distinctly split by a thick epiphyseal bar. Usually confused as representing M. vittatus or M. tengara. Mystus colvillii (Günther, 1874)
See images of type specimens and see further discussion under Mystus pelusius (Solander, 1794). Mystus dibrugarensis (Chaudhuri, 1913)
Mystus gulio (Hamilton, 1822) (Please see image of live specimen)
Mystus horai Jayaram, 1954
See image of a type specimen. It has been included here as it was originally described as a sub species of Mystus vittatus (Bloch, 1794), although according to the original description there are no stripes on the body of M. horai, just a faint black ‘shoulder’ mark. Preserved colouration said to be brownish yellow above, dull grey underneath. Type specimens originate from the Indus River, Kalabagh, Pakistan; the largest specimen being 8.4 cm SL. Mystus keletius (Valenciennes, 1840)
Bagrus albilabris (Specimens
from Pondicherry, and possibly Sri Lanka) Valenciennes,
1840? M. keletius was originally described from Java, and Pondicherry. Ng (2002) examined the type specimens and concluded that the one from Java represents Mystus nigriceps (Valenciennes, 1840); and after a lectotype designation restricting the type locality for M. keletius to Pondicherry [Puducherry], India.See image of the lectotype (MNHN A.9011) from Pondicherry, India; the specimen measures approx. 9cm SL. In earlier versions of this article I said that Valenciennes stated that the colour and pattern was very similar to Mystus tengara (Hamilton, 1822) in its colouration and pattern, but differed due to the supraoccipital and humeral (cleithral) processes being more granulated, the supraoccipital process being longer, the dorsal fin appearing rounder and the maxillary barbels being shorter. This is an error. Valenciennes actually stated that (translated from French) “is a species very close to the preceding, and which is confused at Pondicherry under the same name”. The preceding species was Mystus cavasius (Hamilton, 1822) (although the ‘M. cavasius’ at Pondicherry is actually M. seengtee (Sykes, 1839) – see Chakrabarty & Ng (2005)), and at the end of that account Valenciennes refers to M. tengara of Bengal and that is why my error was made. Ferraris (2007) classed it as a junior synonym of M. vittatus; Pethiyagoda et al, 2008 tentatively agreeing with him. In the earlier versions of this article I said it was different to M. vittatus based on the more rugose bony plates, slightly different fontanel, and shorter barbels. Also, Baensch & Evers (2002) show a picture captioned as M. keletius and state that it has a yellow/silvery colouring, with a “dark shoulder mark”, two silvery to golden strips bordering the lateral line, and black marks near the dorsal and caudal fin. Jayaram & Sanyal (2003) state it is "Brownish turning dull white beneath. A dark shoulder spot and a light band along lateral line present. Dorsal and caudal fin tips tinged black, anterior portions of anal fin black". If this colour and pattern is correct for specimens from the type locality and they are conspecific with the lectotype then M. keletius is valid in my opinion. Jayaram & Sanyal also show a drawing of a specimen which is clearly different in its morphology to that of M. vittatus.Interestingly Easa & Shaji (2003: Fig. 118) picture a specimen that they caption as M. gulio (Hamilton, 1822). The true M. gulio appears (in my opinion) to be restricted to north, and north eastern India, Nepal, and Bangladesh (and possibly Pakistan). The identity of the species distributed in Myanmar, Thailand, Vietnam, Malaysia, and Indonesia has not yet been proved (and there are several names already available if these do prove to be distinct). It is possible that the species captioned as M. gulio and present in south western India, and Sri Lanka is actually M. keletius. If this is the case the syntypes of M. albilabris from Pondicherry (and possibly Sri Lanka – if the type information in Eschmeyer, 2008 is correct – the original description does not mention Sri Lanka) may be synonymous with M. keletius; certainly they have similarly originally described colour patterns, overall shape and proportions, relatively short adipose fins, and similar cranial morphology (see images of some of the MNHN syntypes of M. albilabris on www.fishbase.org). M. keletius and M. albilabris were described in the same work. Because M. albilabris is based on syntypes that may not be conspecific with each other due to their differing localities, it may be an idea in the future to designate one of the specimens from “Ganges” as the lectotype. If it came down to being the first reviser I consider that M. keletius should take precedence over M. albilabris.Based on Valenciennes showing that M. keletius is similar to M. seengtee; the information and drawing in Jayaram & Sanyal; the possibility of it being what has been incorrectly captioned as M. gulio in south western India; and the lectotype of M. keletius being different to the holotype of M. vittatus, M. keletius should remain as a valid species until further specimens can be studied and compared to M. vittatus. Although the pattern is more like M. seengtee it clearly differs from that species in morphology, the obvious difference being in the length of the adipose fin (longer in M. seengtee) and the much longer fontanel of M. seengtee (see images above of a non type specimen, and the possible holotype of M. seengtee). Although there are zoogeographical differences to M. gulio, morphological differences need further work. It differs from the similarly patterned M. oculatus (and therefore M. armatus) by the shape and structure of the fontanel (longer and broader, and split in M. oculatus). See under M. ankutta for differences to that species. Mystus malabaricus
(Jerdon, 1849)
(Please
see images of live specimens) |
 (Click
thumbnails for image)
(Click
thumbnails for image)
|

|

 (Click
thumbnails for image)
(Click
thumbnails for image)
|
 (Click
thumbnails for image)
(Click
thumbnails for image)
|
I wondered if based on the locality and that colour and pattern if it was possible that Jerdon’s species is a synonym of M. armatus and/or M. oculatus, the latter being the senior synonym. However, Jayaram & Sanyal (under the account of M. armatus) state that although it is brownish on top, and becoming lighter beneath, freshly collected specimens have a dark band on the sides along the lateral line ending in a dark blotch at the base of the caudal fin. Also a dark band along the anal fin and upper half of the dorsal fin. This would make it very close to M. dibrugarensis in terms of colour and pattern, although that species has a definite tympanum spot, more vivid black markings (with some in caudal fin), and different zoogeography. I doubt that the specimen from Kenchong River, Myanmar listed in Jayaram & Sanyal (2003) is M. malabaricus, I think it will possibly be M. dibrugarensis or even M. castaneus Ng, 2002 in view of the zoogeography. I am aware of at least 2 new species from Myanmar so it is unlikely that the Myanmar specimen is M. malabaricus. Please see my images of a specimen that is very close (if not identical) to the erroneous identification of M. armatus (= M. malabaricus of Jerdon in my opinion – see under M. oculatus). This specimen of unknown origin is 102mm SL, and stopped growing some time ago therefore I consider it an adult. I consider the AMS and NMW specimens shown here represent M. malabaricus. *See images by Hayath of specimens from the southern states of Kerala, Tamilnadu and Karnataka in the western ghats. Very similar to M. ankutta; see that species account for differences. Similar to M. rufescens, but that has a more slender body; the head shape when viewed from above in M. rufescens is more angular; and the fontanel is more triangular at its front portion in M. rufescens. ? Mystus misrai Anuradha, 1986
See discussion under Mystus pelusius (Solander, 1794). Mystus montanus (Jerdon, 1849)
The original description states “Colour greenish above and on the fins; yellow on the cheeks and beneath” and this was the colour and pattern at about 6 inches. See further discussion under M. dibrugarensis about the colour and pattern in live/fresh specimens from different localities as found by Jayaram & Sanyal (2003). Jerdon stated that he “only found it in the river at Manantoddy in Wynaad” which I think is now called Mananthavady, and the river the Kabini (or Kabbani), but Jayaram & Sanyal found it in other localities in Kerala and Tamil Nadu.At first glance it is also similar to M. pulcher but that species has a much more distinctly round tympanic spot, as well as morphometric differences and zoogeography. Mystus multiradiatus Roberts, 1992 (Please see image of live specimen)
See holotype (CAS 76119) which originates from Prachinburi market, Thailand, and live specimen by Kamphol. Largest type specimen is 12.8 cm SL. The small gap between supra-occipital process and basal bones of dorsal fin spine help visually differentiate this species from the similar M. atrifasciatus. It is also distinguished from the similar M. bleekeri by the much less conspicuous and non-indented fontanel, and the lack of dark tympanic spot. Mystus mysticetus Roberts, 1992 (Please see image of live specimen)
See holotype (CAS 76121) which originates from Nakorn Phanom market, Thailand, and live specimen by Allan James. Largest type specimen is 12.9 cm SL. This species has a humeral semi-ocellus, and the tympanum is darkly pigmented but is not a semi-ocellus. Tips of anal and caudal fin often black. The laterally placed eyes (visible when viewed from above or below), and the combination of small adipose fin, humeral semi-ocellus, and dark tympanum differentiate this from all other species. Mystus oculatus (Valenciennes, 1840) |

 (Click
thumbnails for image)
(Click
thumbnails for image)
|
|
(Click
thumbnails for image)
|
 (Click
thumbnail for image)
(Click
thumbnail for image)
|
Type locality for M. oculatus is “Malabar Coast”, India. This is on the south western coast, covering parts of modern day Karnataka, Kerala, and Tamil Nadu states. Day (1865b:v) classed his “denomination” of Malabar as the coastal region from Mangalore (in Karnataka state) to Cape Comorin [Kanyakumari town] in Tamil Nadu state, and inland into the Western Ghats (a mountain range). This includes Cochin (now known as Kochi), which is the type locality of M. armatus It has been kindly pointed out to me that the historical Malabar region does not cover the large area used by Day (including Cochin) but it is clear that Day’s use of Malabar did and this is important when discussing the species he described or covered. See holotype (MNHN 1195) of M. oculatus. According to the original description by Valenciennes, and Day (1877) there is no stripe present on the body in M. oculatus. The colour/pattern is said to be grey at the top, silvery on the sides, lighter beneath. The reason I have included it here is that Jayaram & Sanyal (2003) state it is dark brown above, yellowish beneath with a dark shoulder spot, a dark spot at the base of the dorsal fin (spine), a dark band along the middle of the dorsal fin, and a dark broad band along the lateral line. I’m not sure if this is from observations of the specimens they used, or from the figure in Day (1877), which in my opinion is just trying to illustrate the lateral line itself, not any band of colour or pattern along it. However, they did have four recently preserved specimens so this may be from them and if so it is possible that the body spot and bar quickly fade in older or less well preserved specimens? However, in Easa & Shaji (2003) and Mercy et al (2007) no stripe is present along the lateral line. In the 2004 and 2006 versions of this article I included M. armatus (Day, 1865) as a valid species. However, I raised the point that M. armatus needed checking against M. oculatus as they might be conspecific. Day described M. armatus as new in 1865a from “rivers and backwaters of Cochin, India”, but did not include any details regarding its colour or pattern, but did thankfully say (amongst other morphological details) that the base of the adipose fin equalled that of the anal fin, the supra-occipital reached the basal bones of the dorsal fin, and the fontanel almost reached the origin of the supra-occipital process (which he showed in a drawing). In 1865b he used the same morphological information but went on to say that the colour and pattern was “Bright leaden silvery, lightest along the sides and with a purplish gloss over the cheeks. A black spot just anterior to the root of the dorsal spine. Fins finely dotted with minute black points.” He then stated that the adipose fin commenced a short distance anterior to that of the anal fin. Unfortunately no figure was published of the whole fish and as yet the type specimens have not been definitely identified. However, in my opinion BMNH 1865.7.17.21 does constitute one of the syntypes (one of a number of type specimens originally used by Day), alternatively it may be a specimen used by Day in 1865b (although they were probably one and the same, even taking into account the slightly different statements about the adipose fin). According to the BMNH website the locality for the specimen is “Malabar”. Looking at the descriptions by Day in 1865, the BMNH specimen certainly seems to match. In particular the bony plates on the head (cranium); the size, shape and placement of the fontanel; the size and length of the supra-occipital process; the dorsal and pectoral fin spine morphology; and the darkish mark near the dorsal fin spine base (which on the BMNH specimen looks like it is on the anterior and median nuchal plates) all match. I have compared the BMNH specimen with the holotype of M. oculatus. Although the MNHN specimen is not as well preserved the two specimens appear to closely match in all these factors. In particular the morphology of the bony plates of the head and nuchal plates are virtually identical. The size, shape, and placement of the fontanel match, and the supra-occipital process matches. Although one could be mislead by Day (1877) into thinking that the maxillary barbels of M. armatus are shorter than that of M. oculatus, in 1865a and b it is clear that the maxillary barbels reach the anal fin (in fact in the original description it states that they reach the caudal fin), as they do in M. oculatus. The main possible difference is the size of the eyes. In M. oculatus at first glance they appear larger, however (using images) in the holotype of M. oculatus the eye goes into the head length 3.4 times, and in the syntype of M. armatus it goes in 3.57 times. This is very close. It is my tentative opinion that based on the locality details and the colour, pattern, and morphology that M. armatus is a junior synonym of M. oculatus. Hopefully an ichthyologist will resolve this problem once they have definitely defined the true M. armatus, although unfortunately even in 2008 this still doesn’t appear to be happening. Thanks to Mark Sabaj Peréz and Kyle Luckenbill I have also been able to compare the BMNH syntype with NMW 45165 (see images above) which has previously been highlighted as a possible syntype of M. armatus. The NMW specimen does not match the original description of M. armatus or of the BMNH syntype. Notable differences are in the fontanel. In the BMNH (syntype) the fontanel is very long and narrow, commencing anterior to the eyes and ending near the base of the supra-occipital process, whereas in the NMW specimen it stops well short of the base of the supra-occipital process and its anterior portion is wider than the posterior portion. In the BMNH syntype it is partially split by an epiphyseal bar near the anterior rim of the eye, whereas in the NMW specimen it is not split at all (although there is a slight indentation in the shape). For these reasons I do not consider NMW 45165 as a syntype of M. armatus. There is no mention of stripes in the original description of M. armatus, but in Day (1877, Plate CI fig. 3.) a drawing is shown of a fish with a mid-lateral stripe terminating in a blotch on the body near the base of the caudal fin, and the adipose fin being much longer than the anal fin base. In my opinion this is probably AMS B.7573, which Ferraris et al. (2000) have correctly stated is not a type specimen of M. armatus. AMS B.7573 is from Canara district and as such could represent M. canarensis or M. malabaricus, thus accounting for the differences in pattern and morphology. Thanks to Shobnom Ferdous I have been sent images of AMS B.7573 (see above). It matches exactly with the drawing and account in Day (1877). In the text in Day (1877) he states that M. armatus doesn’t always show the dark lateral line or dark shoulder mark. Unfortunately AMS B.7573 is not conspecific with M. armatus as originally described by Day in 1865. Bearing in mind that BMNH 1865.7.17.21 does match M. armatus as originally described, if one compares AMS B.7573 with that BMNH syntype one can see that not only is the general shape of the head and body different, but that the fontanels are a completely different shape, but more importantly a different length (bearing in mind the drawing and information about this in the original description of M. armatus). The differences between the BMNH specimen and the NMW specimen are mirrored here; the AMS specimen appearing conspecific with the NMW specimen. In my opinion this misidentification in Day (1877) (using a non type and non conspecific specimen) has led to Jayaram (1954) and subsequently (Ng 2002), Jayaram & Sanyal (2003), Eas & Shaji (2003), Jayaram (2006), and Pethiyagoda et al (2008) considering M. armatus as having a dark stripe along the body. Day’s 1877 account of M. armatus (and therefore Jayaram’s et al) shows the maxillary barbels as only reaching around the ventral (pelvic) fins, whereas as stated above it is clear from the 1865 accounts that they are longer. Also, it is clear that as originally described M. armatus has the dark mark around the base of the dorsal fin spine (also present in M. oculatus), and this is missing in the Day (1877) account and therefore in the subsequent accounts by Jayaram et al. My theory is that the M. armatus as originally described is a synonym of M. oculatus, and that Day’s 1877 account (and therefore all accounts since) of M. armatus are actually M. malabaricus of Jerdon. As an example, compare the cranium of the specimen figured in Pethiyagoda et al (2008: fig. 6a) as representing M. armatus with that of the syntype of M. armatus (BMNH 1865. 7.17.21) and you will see that the fontanels are of a completely different shape, placement, and length. See image of a live specimen under M. malabaricus that appears conspecific with the NMW and AMS specimens, but not the BMNH one. As mentioned previously, a final and definite determination and then examination of a syntype of M. armatus and then a comparison with that of the holotype of M. oculatus, with then a comparison with the specimens identified as M. armatus by Jayaram et al, then in turn a comparison with the type of M. canarensis will resolve this issue one way or another. Until then I consider M. armatus a tentative junior synonym of M. oculatus. Mystus pelusius (Solander, 1794)
Junior synonyms: Bagrus halepensis
Valenciennes, 1840 Roberts (1994) tentatively considered M. colvillii (from the Tigris River at Baghdad, Iraq) and M. misrai (from Lake Antioche, Syria) to be junior synonyms of M. pelusius, although he did note that in the future M. colvillii may prove to be distinct. Thanks to Anthony Troncale of the American Museum of Natural History I have now managed to see the original drawing of M. pelusius from Solander (1794), and the text of the description, which is very basic in terms of colour and pattern information, just saying it was predominantly dark silver. The colour pattern of the alleged junior synonym M. misrai (see image of a type specimen) is (in alcohol): pale yellowish brown with head slightly lighter. No spots or stripes present. The drawing of M. pelusius looks more similar to M. colvillii than it does to M. misrai, although the profile of the dorsal fin is different in M. colvillii than the drawing of M. pelusius. The image here of BMNH BMNH 1955.6.25.1 (a syntype of B. halepensis, and possibly of M. pelusius also) looks very different to M. colvillii. M. colvillii is said to be olivaceous in colour, with three narrow, white, parallel, longitudinal stripes, one along, one above, and one below the lateral line. The type specimens are “9 inches long”. According to Ferraris (2007) BMNH 1955.6.25.1 is a syntype of M. halepensis (a junior synonym of M. pelusius). If you compare the M. halepensis specimen with the syntypes of M. colvillii you will see that the whole body, head, and fin shape is different. In addition the fontanel in M. colvillii appears to extend much further forward anteriorly. Based on these points and the different locality data, I consider that M. colvillii is probably valid. M. misrai appears to be a junior synonym of M. pelusius. Mystus pulcher (Chaudhuri, 1911) (Please see images of live specimens)
Described from four small specimens (6.7cm total length) from near the Yunnan border, upper Myanmar. See image of a type specimen, and a live specimen by Kamphol. Described as “Dorsal and upper part of the body dark brown, with lighter or paler whitish brown stripes: one median, from the tip of the snout to the base of the dorsal spine, and two lateral longitudinal on each side, one above and the other below the middle line, which is distinguished by being dotted black for the openings of the lateral organs.” “…nasal and maxillary barbells blackish brown, adipose fin dark brown, dorsal, anal and caudal fins are brownish with black spots on the membranes between the rays.” It has an intensely black tympanic spot/semi-ocellus, and also a spot near the caudal peduncle, followed by a thin white band. Its supraoccipital process meets the basal bones of the dorsal fin. The difference in length of the supra-occipital process differentiates it from M. canarensis. The light body stripes differentiate it from M. dibrugarensis. Differs from M. rufescens by the fact that its fontanel is not split by an epiphyseal bar, and it has a shorter adipose fin. Mystus rhegma Fowler, 1935 (Please see image of live specimen) |
 (Click
thumbnails for image)
(Click
thumbnails for image) |
|
See drawing and image of holotype ANSP 61748 which originates from Bangkok, Thailand, and measures 4.96cm SL; and a live specimen by Kamphol. Roberts (1994) lists specimens up to 10.6cm SL. As per original description: “Very light or pale brown, lower or under surfaces more or less whitish. Upper surface of head and back sprinkled with dark grey dots. Band of dark dots along lateral line and broader one along lower side of trunk and tail parallel. Iris greyish, also maxillary barbell, other barbells whitish. Outer edge of adipose fin dusted with dark grey dots. Caudal dark grey. Other fins pale or whitish.” This is a small and graceful looking species, which reminds me of species of the South American pimelodid genus Pimelodella Eigenmann & Eigenmann, 1888. The lack of any semi-ocellus, the thin dark band across the lateral line, and large gap between the small supraoccipital process and basal bone of the dorsal fin spine differentiate this species from all the others. Mystus rufescens (Vinciguerra, 1890) (Please see images of live specimens) |
 (Click
thumbnails for image)
(Click
thumbnails for image) |
 (Click
thumbnails for image)
(Click
thumbnails for image) |

 (Click
thumbnails for image)
(Click
thumbnails for image) |
 (Click
thumbnail for image)
(Click
thumbnail for image) |
Vinciguerra described the colour and pattern as body tawny reddish, with one spot near the humeral region and one on the caudal peduncle. If one looks at the figure of the holotype there appears to be a thin black stripe along the lateral line. Differs from the similar M. canarensis, M. dibrugarensis and M. pulcher by having its fontanel split by an epiphyseal bar. Mystus vittatus
(Bloch, 1794) |

 (Click
thumbnails for image)
(Click
thumbnails for image) |
 (Click
thumbnails for image)
(Click
thumbnails for image) |

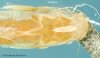
 (Click
thumbnails for image)
(Click
thumbnails for image) |

 (Click
thumbnails for image)
(Click
thumbnails for image) |
Bloch described the colour and pattern as head, ‘back’ fins, and caudal chestnut brown. Other fins steel coloured. Stripes light blue, with yellow interspaces. The plate of the holotype shows the general shape of the body and fins but the accuracy of its pattern is doubtful. See images of the holotype of M. vittatus: ZMB 2939 that originates from Tranquebar, Tamil Nadu, India.Since its description in 1822 from “Ponds of India”, M. tengara has been discussed as being very similar to or perhaps a junior synonym of M. vittatus. Jayaram & Sanyal (2003) have classed them both as valid but they did not have access to the holotype of M. vittatus and as such they may have been using non type specimens and information from years of wrongly identified specimens which has just added to the confusion. They may have used misidentified specimens of M. carcio as representing M. tengara, and therefore incorrectly distinguishing M. vittatus as distinct from M. tengara. If one looks at the original drawings of M. tengara (see images) and compares them with the images of the holotype of M. vittatus, you will see that the structure of the fontanel and supraoccipital process are very similar, the only difference being that in the drawing of M. tengara the front portion of the fontanel is thin, the second is wider, whereas this is the opposite in the holotype of M. vittatus. This could be a mistake in the drawing. Everything else from the description of M. tengara appears to fit M. vittatus. If one looks at the images of ANSP 85780 from Bombay [Mumbai], India (which definitely represent M. vittatus in view of the exact same fontanel and supra-occipital morphology when compared to the holotype) you will see that the pattern, and shape of the body, especially relating to the high back and adipose fin matches that of the drawing of M. tengara. The adipose fin on the holotype of M. vittatus has shrunk in size due to its age. The only possible evidence I have seen so far for classing M. tengara as possibly valid is that of specimen CAS-SU 34858 from Calcutta [Kolkata], India (see images). If you look at the lateral image of this specimen it appears visually very similar to that of ANSP 85780. But if you compare the fontanels of each specimen you will see that the fontanel of the CAS specimen is long and evenly portioned, unlike that of the holotype of M. vittatus and of the ANSP specimen. I feel that for the validity of M. tengara to be properly determined, future ichthyologists need to bear in mind that validity and identity of M. carcio when comparing lots of specimens from different localities. Until then, I feel that M. tengara should be classed as a questionable synonym of M. vittatus. I say questionable because of the possible aforementioned differences, and because M. vittatus comes from southern India, M. tengara possibly representing the Ganges population (but with M. carcio distinct from both). M. vittatus will reach
at least 8 to 9cm SL. |
Acknowledgements |
Thanks to Shobnom Ferdous of Auburn University, USA; Jon Fong of the California Academy of Sciences, California, USA; Mark Sabaj and Kyle Luckenbill of the Academy of Natural Sciences, Philadelphia, USA; Mark McGrouther & Sally Reader, and Mark Allen (formerly) of the Australian Museum, Sydney, Australia; Rémi Ksas of the Muséum National d’Histoire Naturelle, Paris, France; Dr Peter Bartsch of the Institut fuer Systematische Zoologie, Museum fuer Naturkunde der Humboldt-Universitaet zu Berlin, Helmut Wellendorf, Naturhistorisches Museum Wien, Austria for the provision of the images of preserved specimens. Rohan Pethiyagoda for his opinions and information. Anthony Troncale of the American Museum of Natural History, New York, USA for the drawing of M. pelusius. Rahul Kumar for permission to use his images; Kamphol Udomritthiruj of AquariCorp for allowing use of his images and for his opinions; Rohan Pethiyagoda for the image and description of M. ankutta, Allan James for the use of his images and his efforts to post this on ScotCat; Roy Blackburn for the opportunity to photograph his fish. |
References |
Anuradha,
S., 1986. Contributions to the study of bagrid
fishes. 19. Systematic position of Macrones halepensis
colvillii Hora & Misra, 1943, with description of
a new species (Siluriformes, Bagridae). Rev. Suisse
Zool. v. 93 (no. 2): 291-296. |
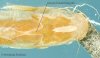
 (Click
thumbnail for image)
(Click
thumbnail for image)
































If you would like to contribute an article, please e-mail me. You will of course be credited for your work.
If you would like to donate any denomination of money to the site just click the above link button. All proceeds will go to running the site and hopefully to keep it going for a few years yet.
Print or e-mail this article below
